Lecture 27:
The structure of plant communities I
Basic questions:
-
Why are certain structures of vegetation restricted to certain combinations
of climate and soil?
-
What determines the number of different species in a given vegetation?
-
What determines the relative abundance of different species in a given
vegetation?
-
What determines the observed gradients in species composition on a geographical
scale or along a mountain slope?
-
What factors influence the succession of species after disturbance?
-
What makes different species coexist in the first place?
Individuals, populations and communities:
-
Individual
: not all individuals can be treated as one if their survival, growth
or fecundity are different; we then must distinguish
between individuals depending on their stage (e.g. seeds, vegetative
rosettes, flowering plants), size or age
(see Lecture 26)
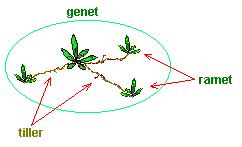
: break-down of the concept of the 'individual' in clonal plants
.: genet (derived from a single zygote)
.: ramet (the potentially physiologically independent module of a genet)
: break-down of the concept of 'individual' in sexually reproducing plants
with genetic mosaic (i.e., the occurrence of
branches with different genotypes on the same plant as a consequence
of somatic mutations)
.: significance of natural selection between branches with different
genotypes within a plant
-
Population
: all individuals of a given species occupying an area which an ecologist
has circumscribed for the purpose of study
.: in this definition there need be no interaction or relation between
the individuals other than that they happen to
be together in the same place at the same time
: in a more strict definition definition we may require in addition that
all individuals (potentially) interact with one another
either directly (pairwise contest among neighbors: shading,
allelopathy, etc.) or indirectly (e.g., plant A
competes with plant B, who competes with plant C, so that
A indirectly affects C even though they are not
neighbors)
.: It could be argued that exponentially increasing or declining populations
without density- regulation are not real
populations in the stricter sense.
: significance of spatial scales
.: seed and pollen dispersal distances
.: closed populations or open populations with immigration and emigration
: significance of time scales
.: infrequent long distance dispersal
.: increasing time-scale is likely to increase the spatial scale of
what we would consider a population as well,
because on a longer time scale we include the more rare
events of very large distance dispersal
: meta-population ("population of populations")
-
Community
: all individuals of a given set of species occupying an area which
an ecologist has circumscribed for the purpose of study
.: in this definition there need not be any relation or interaction
between the different species in the community.
They could be populations of non-interacting species that just
happen to be in the same place at the same time
: in a more strict definition we may require that all individuals (potentially)
interact with one another either directly or
indirectly, and both within species and across species borders
: community structure: a list of species and their relative abundance
and spatial distribution in a community
: significance of spatial scales
: significance of time scales
-
So, what is a population/community after all?
: depends on the spatial scale and time-scale that is significant for
the question under investigation
: increasing spatial scale and time scale:
.: descriptive (which species occur in what type of environment and
in combination with what other species?)
.: plant community dynamics
.: evolutionary ecology
Clements' view of communities
-
community as an integrated dynamical system of interacting species
-
between-species interactions essential for community structure and
development
-
succession: change in community structure resulting from the non-seasonal,
directional and continuous pattern of colonization and extinction;
primary succession: soil-forming process, immigration and colonization,
no soil seed bank present yet;
secondary succession (after a disturbance): minimal changes
in soil, recruitment from soil seed bank dominates immigration;
climax: the presumed endpoint of a succession; a community that
has reached a steady state
-
predictable succession; single climax dominates in any given climatic
region, independently of starting point (sand-dune or abandoned old field
etc.)
Gleason's view of communities
-
communities as a collection of coexisting species; between-species
interactions unimprotant for determining community structure
-
community structure as a consequence of differences in dispersal capability
of plants, proximity of source populations, and environmental selection
-
succession strongly history-dependent (which species happens to come along
next?); no fixed climax
The niche concept
-
"a multi-dimensional description of a species' needs, habitat requirements
and environmental tolerances" (Hutchinson, 1957), or "the limits, for all
important environmental features, within which individuals of a species
can survive, grow and reproduce" (Begon et al, 1996)
-
fundamental niche vs realized niche (cf. physiological optimum and ecological
optimum)
-
niche width; generalists and specialists; niche overlap
-
use of niche concept in explaining species distributions, species richness
Coexistence and species richness (three principles)
-
Principle 1: competitive exclusion principle (Gauses principle)
: two species with the same niche cannot coexist
: ==> necessity of niche differentiation
: species richness and
.: niche width
.: niche overlap
.: length of niche axis
: implementation problems
.: fundamental niche or realized niche
.: tautology: if two species coexist they apparently must have different
niches, but as long as we cannot determine
a species' niche in some independent way, this doesn't
really help us understand why coexistence occurs
-
Principle 2: intra- vs inter-specific competition
: two species can coexist if for both species the intra specific competition
is greater than the inter-specific competition
: advantage: you can actually measure whether intra specific competition
is more important than inra-specific
competition in a given species mixture, e.g. by comparing the
yield of the plant labeled * in the following two situations
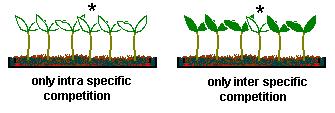
: still, doesnt explain why coexistence is possible
-
Principle 3: frequency-dependent growth rates
: we've seen that density-dependence is necessary for a species to
persist for any reasonably long period of time (see
lecture 25)
.: can density-dependence also promote species coexistence? Answer:
NO.
.: with density-dependence (and monotonically decreasing growth rate
as density increases) we get that the species with
the highest density at equilibrium in a mono-culture can
exclude all others
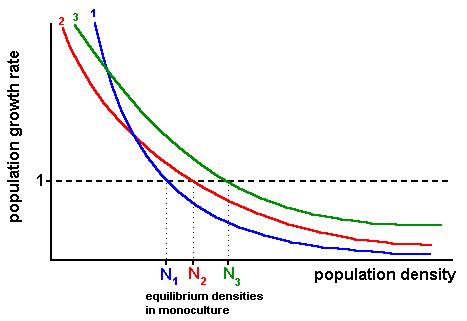
- a monoculture of species 1 at its equilibrium can be
invaded by species 2 and 3 if introduced at initially low density,
because both have a population growth
rate greater than one.
- a monoculture of species 2 at its equilibrium can be
invaded by species 3, but not by species 1
- a monoculture of species 3 cannot be invaded by any
of the others
: frequency-depend population growth rates
.: growth rate does not depend on total density, but on the relative
frequencies of the various species present
.: "advantage of the rare species": a species has a high population
growth rate when rare, but a low growth rate
when common (e.g., due to frequency-dependent herbivory
or other reasons; see next lecture)
.: protected coexistence
Unification of the three principles
-
Intra- and inter-specific competition
: when a species is rare, it experiences mainly inter-specific
competition
: when a species is common, it experiences mainly intra specific
competition
: if (intra specific comp.) > (inter-specific comp.),
then it follows that the rare species has a relative advantage over the
common species
: In this way we have translated principle #2 to principle #3
: high intra specific competition and low inter specific competition
is only one way to get frequency-dependent growth
rates and advantage of the rare species (see later for other
examples)
-
Competitive exclusion and niche overlap
: if two species have only little niche overlap, then inter-specific
competition is likely to be smaller than intra
specific competition
: the larger the niche overlap, the larger the inter-specific
competition, and the less likely that the two species can
coexist
: In this way we have translated principle #1 to principle #2, which
in turn we already had translated into principle #3
: reducing niche overlap is only one way to reduce inter specific
competition; there are other ways too (see later for
other examples)
-
Conclusion:
: principle #3 is the most general principle of the three.
Mechanisms for species coexistence
-
stable environments
: small-scale spatial heterogeneity
.: different types of microhabitat (or safe-sites)
.: competitive exclusion within each microhabitat
.: Gauses principle on a small spatial scale
.: specialists vs generalists?
: competitive ability/dispersal trade-off
.: one type of safe-site => both species have the same (fundamental)
niche
.: random seed dispersal
.: the competitive inferior species exploits the safe-sites that by
chance remain unoccupied by seeds of the more competitive species, and
is able to do this because of its own superior dispersal capabilities.
.: e.g. seed size/seed number trade-off
.: advantage of the rare species
.: number of species that could coexist in this way is limited by the
degree of competitive asymmetry
: early germination/survival trade-off
.: competitive advantage of early germination
.: lower density-inependent mortality for later germination during
a more favorable time during the season (no night frost, e.g.)
.: one type of safe-site => both species have the same (fundamental)
niche
.: random seed dispersal
.: the species with late germination exploits safe-sites that by chance
remain unoccupied with seeds of its superior competitor with early germinating
seeds. The competitive disadvantage is offset by a lower density-independent
mortality by germinating at a more favorable time during the season
.: advantage of the rare species
.: number of species that could coexist in this way is limited by the
degree of competitive asymmetry
: aggregated distributions
.: aggregation => strong sib-competition
.: intra specific comp. > inter-specific comp.
: frequency-dependent herbivores or seed predators
.: plant-switching by a generalist
.: each species has its own specialist that prevents
.: advantage of the rare species
-
fluctuating environments
: niche differentiation in time
.: good and bad years different for two species
.: survival during bad years (storage effect) as seeds, rosettes,
underground etc.
.: growth during good years must
:: compensate for loss during bad years, and
:: be sufficient to grow away from extinction boundary
.: niche differentiation in time makes intra specific competition greater
than inter-specific competition
: high-risk/high-quality sites and low-risk/low-quality sites
.: two types of site: stable and unstable
.: two species:
:: disperser exploits high-risk sites and uses low risk sites as storage/source,
and
:: non-disperser exploiting low-risk sites
.: difference in site quality due to differences in density (high-risk
sites are regularly emptied by a disaster)
.: difference in local competitive ability due to differences in seed
losses due to dispersal
-
refuges (Principle #4?)
: in space
.: different microhabitats; tollerance of different extreme conditions
.: aggregation or limited dispersibility of competitively superior
species creates competitor-free space for the competitively inferior species
: in time
.: separation of growth in time during the season (not realy coexistence)
.: storage effect
.: high risk/high quality and low risk/low quality sites