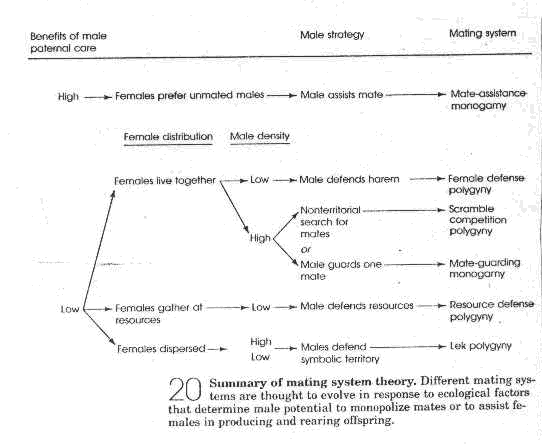
Determinants of Mating Systems
II. Observed patterns
A. Monogamy - 1 female / male
1. > 90% of birds, females disperse farther presumably to pick best resource
2. < 10% of mammals (e.g. canids, beaver, carnivorous bats)
a. Males help provision young - canids, carnivorous bats, marmosets
b. Females solitary and males can only defend 1 female - dik-dik, some rodents
3. very rare in insects (carrion beetles), fishes and herps
B. Polygyny - >1 female / male
1. > 90% of mammals, males defend females when they can, males disperse farther
presumably to find a female group
a. Females solitary, range of 2 or more defensible by male - prosimian primates
b. Females solitary, range not defensible - moose, orangutan
c. Females social, range defensible by male
i. seasonal harems - elephant seals, red deer
ii. permanent harems - hamadryas baboons, spear-nosed bats, zebra
d. Females social, range not defensible
i. Female movements predictable, males wait for females on territories
e.g. topi, waterbuck, Uganda kob - analogous to bird leks
ii. Female movements unpredictable, males follow estrous females
e.g. elephant, moutain sheep
2. In polygynous birds, males defend food or nesting sites
(blackbirds, grouse, cotingas)
c. Polyandry - > 1male / female
1. Rare in birds, very rare in mammals, but fairly common in fishes
2. This occurs in birds where resources are very patchy and available for only a
short period.
a. Sequential - sanderling, Temminck's stint, spotted sandpiper
b. Simultaneous - phalarope, jacana
c. Cooperative - pukeko
d. Promiscuity - both sexes mate multiple times
1. Common in insects
2. Hibernating bats ??
III. Ecological factors influencing mating systems
A. Unequal resource (food or nesting sites) partitioning by males caused by
1. Spatial habitat heterogeneity and fixed territory size - the polygyny threshold model
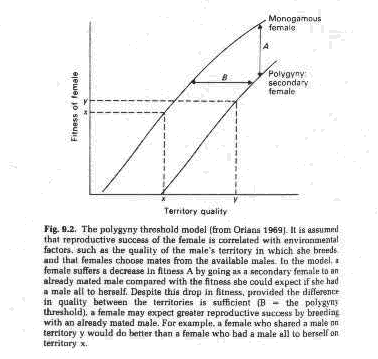
Predictions of the polygyny threshold model:
a. Polygyny should be most common in patchy habitats
b. Male territory quality influences number of females attracted
c. Females who mate with already mated males should have at least as high reproductive success as females choosing unmated males
d. Blackbird experiments show
i. Higher fledging success in larger harems
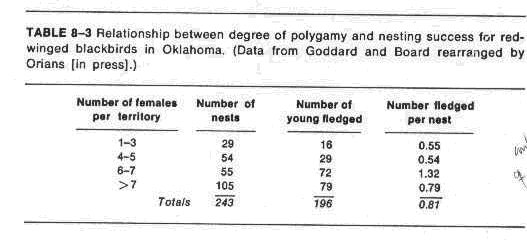
ii. Harem size correlates with habitat quality
iii. Female removals do not lower fledging success; therefore, females do not pay any cost to being secondarily polygynous
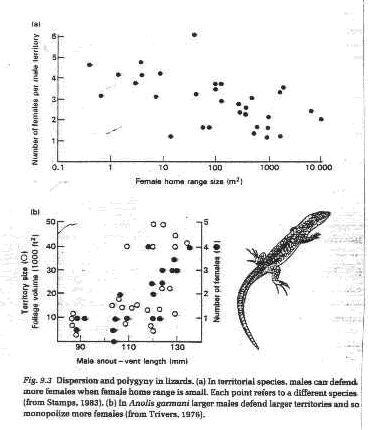
2. Spatial habitat homogeneity and
a. Variation in male size or age which affect male territory size, e.g. lizards
b.Asynchrony of settlement
c. Male error in evaluating resource quality
d. Site fidelity between years and temporal resource variation
B. Female movement predictability
1. If females require a resource, males may wait for them there (resource defense polygyny)
2. If females move through a region, males may defend traditional sites (lek)
3. If females are receptive in synchrony, males cannot monopolize several unless they are sedentary (elephant seals)
C. Female sociality
1. Group living species facilitate male defense (female defense polygyny)
2. Solitary species - males often defend territories that overlap several female territories.
D. Length of breeding season
1. Long breeding seasons permit sequential polygamy
2. Rich resource and short breeding seasons allow polyandry
IV. Sexual conflict
A. Evidence
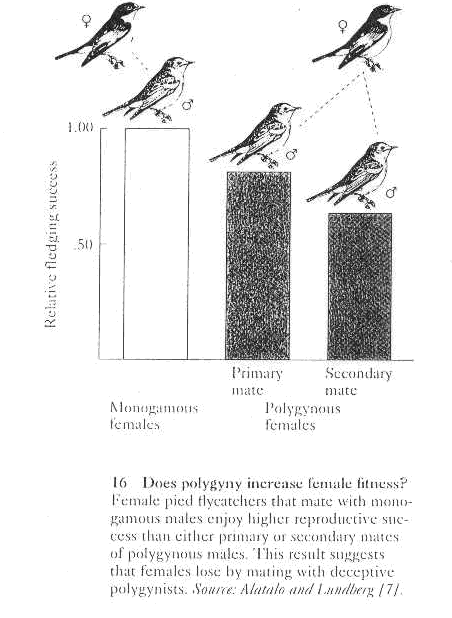
1. Secondary females pay a cost of polygyny in pied flycatchers
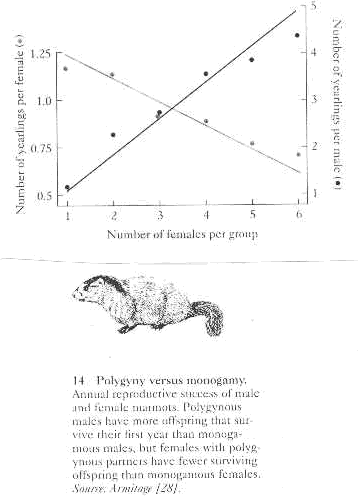
2. Yellow-bellied marmots exhibit decreased female productivity with increased polygyny
B. General expectations
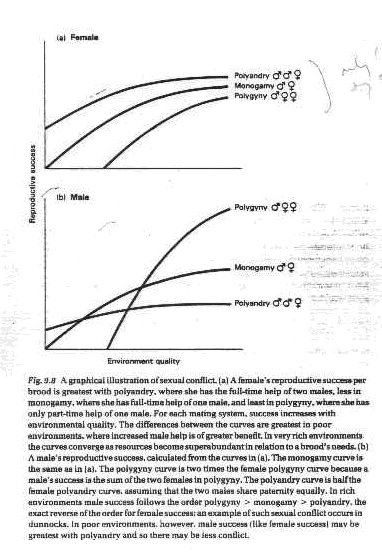
1. Good quality environments
a. Female repro. success: polyandry > monogamy > polygyny
b. Male repro. success: polygyny > monogamy > polyandry
2. Poor quality environments
a. Female repro. success: polyandry > monogamy > polygyny, but differences are greater than in good environments
b. Male repro. success can be polyandry > monogamy > polygyny
C. Dunnocks - mating system changes with access to resources
1. Polygyny occurs in small territories, polyandry in large territories
2. Provisioning caused a reduction in female territory size and a change from polyandry (or polygynandry) to monogamy (or polygyny)
3. Consequently, females have higher repro success in poor environments while successful males have higher repro success in rich environments
4. Females solicit copulations from males to keep them around
5. Male feeding rates are proportional to their paternity
D. Multiple mating by females is common, but not yet well understood. Could be due
to
1. Material benefits, e.g. parental care in dunnocks
2. Genetic benefits, e.g. pseudoscorpions (perhaps!)

V. Factors favoring male aggregations on leks
A. Defining criteria - often suggested as a default mating system when males can't
defend resources or females - only benefit to females from males are genes
1. No paternal care
2. Males cluster on traditional display sites
3. Mating aggregations occur away from any resource required by females
4. Females are free to choose between displaying males
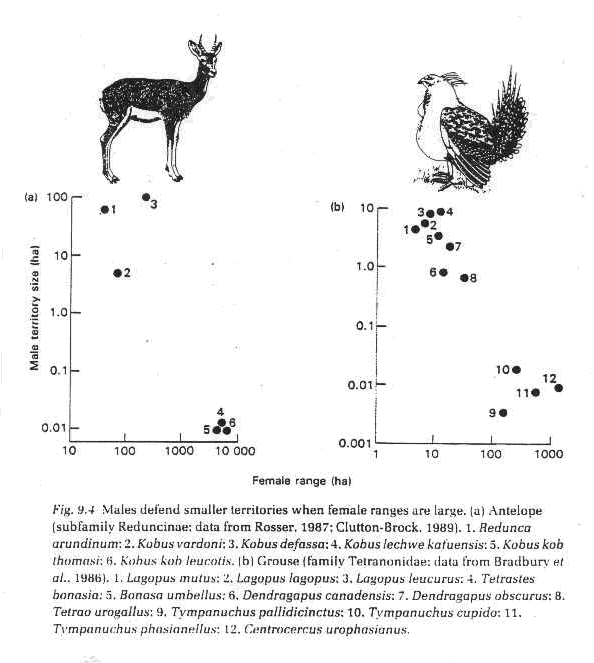
B. Female home range increases as male territory decreases and mating system switches from resource defense to lekking in antelope and grouse
C. Explanations involving male benefits:
1. Conspicuousness - males cluster to increase signal range or time of signal emission
(unlikely)
2. Males require specific display habitats which are limiting (never proven)
3. Hotspot model - males cluster at sites where female home ranges overlap
a. Males sample sites and then use ideal free distribution rules
b. Settlement at a site devalues that site for future settlers and for others that
might encounter females elsewhere in their range
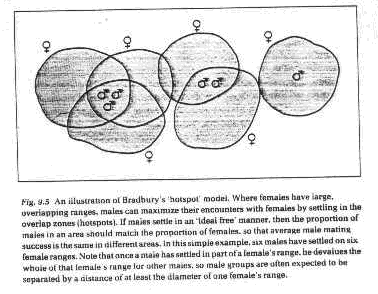
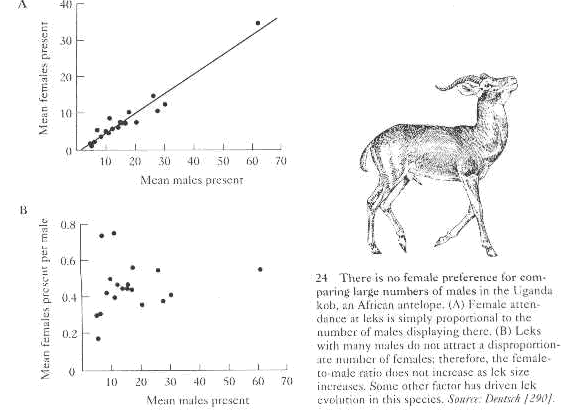
c. Supporting evidence
i. Equal male mating success on Uganda Kob leks
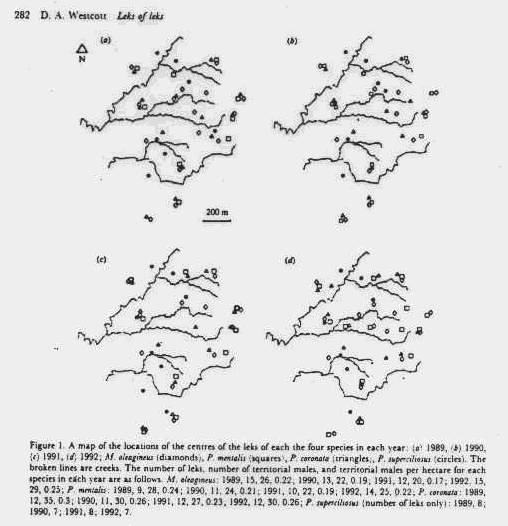
ii. Leks of multiple species sometimes co-occur, consistent with placement where female encounter rate is maximized
d. Negative evidence - female density does not correlate with lek formation in Kafue lechwe (?)
D. Explanations involving female benefits:
1. Direct benefits
a. Mating in an aggregation reduces predation on females (unlikely)
b. Mating in less desirable habitats reduces competition for resources with females
c. Females have a better chance to identify males with "good genes" - often
referred to as hotshots
i. because they have a larger n from which to choose best males
ii. because they can copy each other to ensure unanimity in choice
iii. Supporting evidence is that females clearly choose particular males on leks
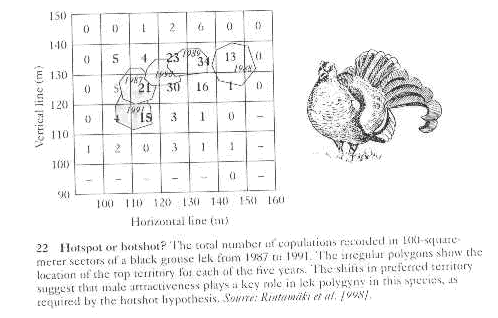
2. Indirect benefits
a. Females respond to "supernormal" stimulus of several males
b. An arbitrary female preference for larger (or smaller) leks can lead to lek evolution
by a Fisher type runaway process
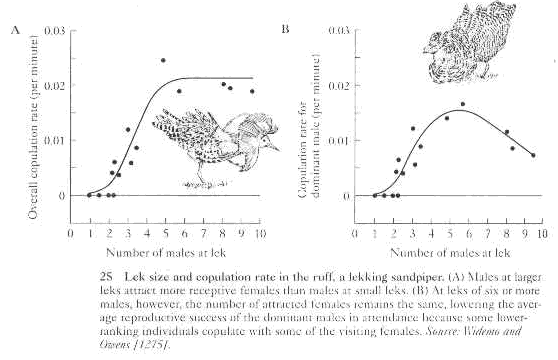
D. Potential cues females use on leks to choose a mate
1. Position - no direct evidence
2. Dominance - no cues predict linear dominance hierarchy, when those exist must
be due to some conventional rule
3. Phenotypic cues
a. Morphological
b. Behavioral - vigor (males can't cheat)
References
Bradbury, J. W. & R. M. Gibson 1983 Leks and mate choice. In: Mate Choice. (P.
P. G. Bateson, ed.) Cambridge University Press, Cambridge.
Bradbury, J., R. Gibson & I. M. Tsai 1986 Hotspots and the dispersion of leks.
Anim. Behav. 34: 1694-1709.
Choe, J.C. & B.J. Crespi (eds) 1997 Mating Systems in Insects and Arachnids.
Cambridge University Press, Cambridge.
Clutton-Brock, T.H. and P.H. Harvey 1978 Mammals, resources and reproductive strategies.
Nature 273:191-195.
Davies, N.B. 1991 Mating systems and ecology. In: Behavioral Ecology, an Evolutionary
Approach. 3rd ed., Sinauer, Sunderland, pp. 263-294.
Gibson, R.M. 1996 A re-evaluation of hotspot settlement in lekking sage grouse. Anim.
Behav. 52:993-1005.
Höglund, J., R.V. Alatalo, R.M. Gibson & A. Lundberg 1995 Mate-choice copying
in black grouse. Anim. Behav. 49:1627-1633.
Hugie, D.M. & D.B. Lank 1997 The resident's dilemma: a female choice model for
the evolution of alternative mating strategies in lekking male ruffs (Philomachus
pugnax). Behav. Ecol. 8:218-225.
Lank, D. B. & C. M. Smith 1987 Conditional lekking in ruff (Philomachus pugnax).
Behav. Ecol. Sociobiol. 20: 137-145.
Parker, G. A. 1978 Evolution of competitive mate searching. Ann. Rev. Entomol. 23:
173-196.
Queller, D. C. 1987 The evolution of leks through female choice. Anim. Behav. 35:
1424-1432.
Searcy, W.A. and K. Yasukawa 1989 Alternative models of territorial polygyny in birds.
Amer. Natur. 134:323-343.
Westcott, D.A. 1994 Leks of leks: a role for hotspots in lek evolution? Proc. R.
Soc. Lond. B 258:281-286
Westcott, D.A. 1997 Lek locations and patterns of female movement and distribution
in a Neotropical frugivorous bird. Anim. Behav. 53:235-247.