Evolution of Eusociality
I. Requirements and distribution of eusociality
A. Requirements
1. Overlapping generations
2. Reproductive division of labor including nonreproductive (or sterile) workers
3. Parental care in a permanent nest
B. Distribution
1. Hymenoptera
a. Ants - all species, although there is a parthenogenetic species with no queens! Many show evidence of caste polymorphism - different morphological forms perform different tasks within the colony.
b. Bees - some, may have evolved multiple times
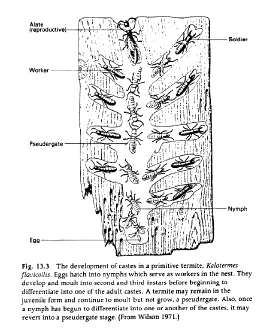
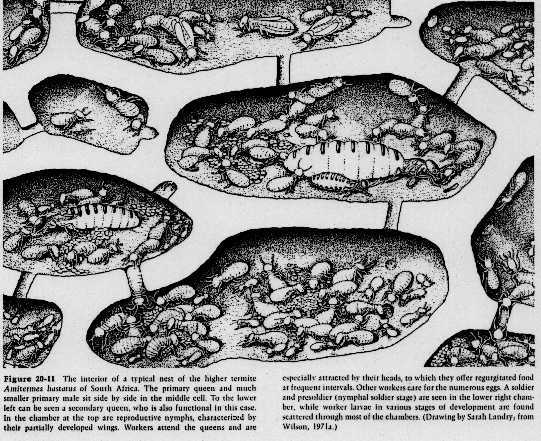
c. Wasps - only in Vespids, variable expression2. Termites - all species are eusocial with cellulose-digesting bacteria, and exhibit caste polymorphisms
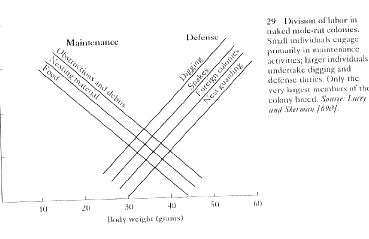
3. Naked mole-rats - any worker can eventually reproduce, food is advertised with calls and chemical trails
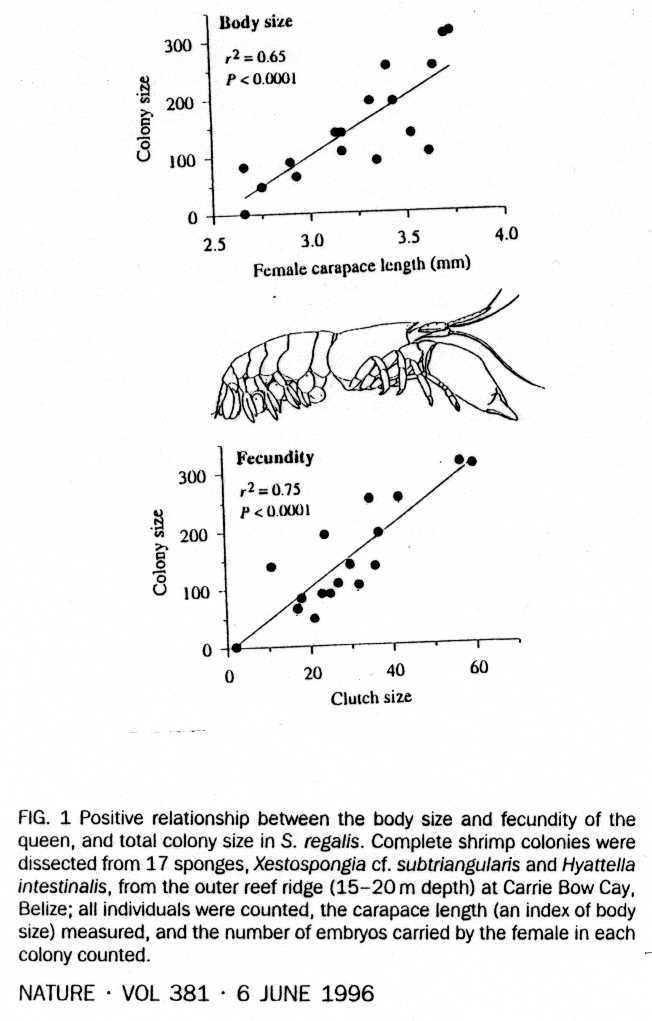
4. Caribbearn shrimp in sponges - colonies of up to 300 shrimp in one sponge, single reproductive pair, larger workers defend sponge against other shrimp
5. Spiders - no caste polymorphism, group living on a common web has evolved multiple times, sterility not yet demonstrated
6. Aphids - a few species, have soldier forms, and are periodically asexual
7. Beetle - one Australian species that builds colonial burrows in eucalyptus trees
II. Possible explanations for worker sterility
A. Kin selection
1. Because of haplodiploidy, workers will be more closely related to their sisters (r = 3/4) than to their own offspring (r = 1/2) if their mothers mated only once.
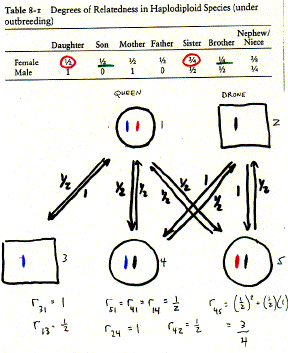
2. But, they are more closely related to their own male offspring (r = 1/2) and their nephews (r = 3/8) than their brothers (r = 1/4). Therefore, expect workers to lay unfertilized eggs. Example: bumblebees and stingless bees are monandrous and have high levels of worker reproduction.
3. If mothers are polyandrous (mate multiple times), then workers may be more closely related to their brothers than to half-nephews (r=1/8). Expect workers to kill unfertilized eggs laid by other workers. Example: honeybees and yellowjackets are polyandrous and have low levels of worker reproduction.4. What about diploid eusocial animals (e.g. termites, naked mole rats)?
a. One proposed hypothesis is that these populaitons undergo cycles of inbreeding. With high levels of inbreeding-mother-son and sister brother can rapid approach r>3/4 for both males and females.
b. But high levels of inbreeding can lead to inbreeding depression: lower fitness due to increased frequency and exposure of deleterious mutations. Thus inbreeding might alternate with some dispersal. A rare disperser morph is found in mole rats: it is fatter, attempts to disperse in captive settings, solicits matings with non-colony members. Once settled reverts to xenophobia and loses fat stores.
B. Parental manipulation (originally viewed as different from kin selection, but most attempts to test it have invoked kin selection, e.g. sex ratios)
1. If parents can prevent their young from reproducing, then it may be better to help.
2. A mother that traded her eggs with her daughters would trade grandkids for offspring and gain a two-fold reproductive advantage. The daughter trades offspring for siblings, and assuming a 1:1 sex ratio, loses nothing and should be less motivated to resist exploitation unless the queen mated multiple times causing sisters to be less closely related than 0.5.
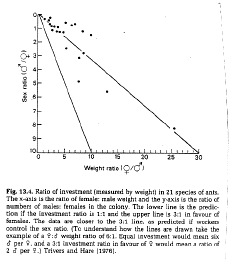
3. Trivers and Hare proposed tested this idea by examining coloy sex ratios of reproductives. They found 3:1 investment ratio in females among monoygynous species which is what would be predicted if the offspring control reproduction (i.e. mating value of a male x relatedness to brothers = mating value of a female x relatedness to females: 3 x 1/4 = 1 x 3/4). Polygynous species and slave-making ants showed 1:1 sex ratios as would be predicted.
4. Mueller confirmed this result with an experimental test in which he removed queens from a monogynous eusocial bee. When queens were removed, a daughter took over reproduction removing the asymmetry in relatedness (her sisters are helping now to produce nieces and nephews, both related to the sisters by 3/8, rather than to rear siters with r=0.75 and brothers with r=0.25). The daughter queen colonies did produce more males (63%) than unmanipulated queen colonies (43%).
III. Routes to sociality
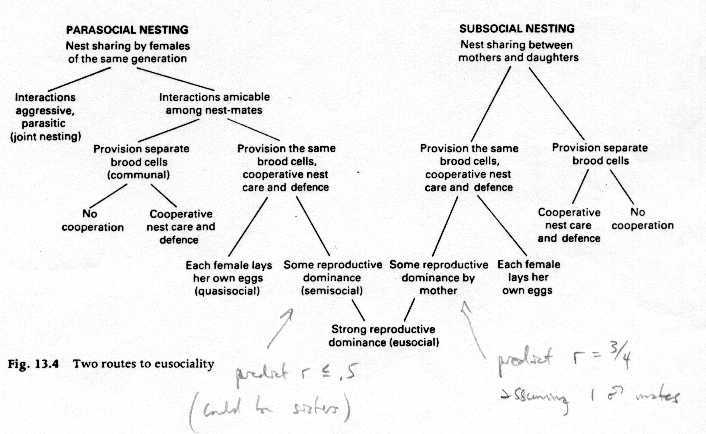
A. Subsocial beginning: single females with offspring remaining to help
1. Single foundress colonies such as honey bees, some ants, e.g. Solenopsis, and wasps
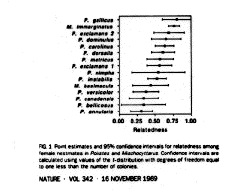
2. Predict high levels of relatedness, which occurs in some species, although in some paper wasps relatedness varies due to multiple matings by queens or multiple females reproducing
B. Parasocial beginning: joint-nest founding
1. Semisocial when some foundresses stop reproducing and just help
2. Some polistine wasps, Harvester ants, some termites
3. Predict low levels of relatedness, e.g. in swarm-founding wasps r ~= 0.2

4. Requires some other reason for eusociality than kin selection, e.g. mutualism over founding a nest
5. Interestingly, recent evidence indicates less aggression occurs when joint foundresses are unrelated (harvester ants) than related (polistine wasps).
C. Ecological factors impose mutualism
1. Nests may be essential resource that is very costly to build, maintain, or defend. Cost of building own nest may outweigh cost of staying in established nest.
2. Nest success depends on the number of foundresses and colony size (as in eusocial shrimp). Benefit of staying versus leaving increases as more animals stay.
Aoki, S. 1977 Colophina clematis (Homoptera: Pemphigidae), an aphid species with 'soldiers.' Kontyu, Tokyo 45:276-282.
Atkinson, L. & E.S. Adams 1997 The origins and relatedness of multiple reproductives in colonies of the termit Nasutitermes corniger. Proc. R. Soc. Lond. B 264:1131-1136.
Bartz, S. H. 1979 Evolution of eusociality in termites. Proc. Natl. Acad. Sci. USA 76:5764-5768.
Benton, T.G. and W.A. Foster 1992 Altruistic housekeeping in a social aphid. Proc. Roy. Soc. Lond. B. 247:199-202.
Bourke, A.F.G. and N.R. Franks 1995 Social Evolution in Ants. Princeton University Press, Princeton, N.J.
Duffy, J. E. l996. Eusociality in a coral-reef shrimp. Nature 38l:5l2-5l4.
Grafen, A. 1986 Split sex ratios and the evolutionary origins of eusociality. J. Theor. Biol. 122:95-121.
Hamilton, W. D. 1964 The genetical evolution of social behavior. J. Theor. Biol. 7:1-52.
Hamilton, W.D. 1972 Altruism and related phenomena, mainly in the social insects. Ann. Rev. Ecol. Syst. 3:193-232.
Jarvis, J. U. M. 1981 Eusociality in a mammal: cooperative breeding in naked mole-rat colonies. Science 212:571-573.
Jarvis, J.U.M., M.J. O'Riain, N.C. Bennett & P.W. Sherman 1994 Mammalian eusociality: a family affair. TREE 9:47-51.
Keller, L. 1993 Queen Number and Sociality in Insects. Oxford University Press, New York.
Kent, D.S. and J. A. Simpson 1992 Eusociality in the beetle Australoplatypus incompertus (Coleoptera: Curculionidae). Naturwissenschaften 79:86-87.
Michener, C.D. and D.J. Brothers 1974 Were workers of eusocial Hymenopera initially altruistic or oppressed? Proc. Natl. Acad. Sci. 71:671-674.
Mueller, U.G. 1991 Haplodiploidy and the evolution of facultative sex ratios in a primatively eusocial bee. Science 254: 442-444.
Queller, D.C., Strassmann, J.E. & C.R. Hughes 1988 Genetic relatedness in colonies of tropical wasps with multiple queens. Science 242:1155-1157.
Reeve, H.K. 1995 Partitioning of reproduction in mother-daughter versus sibling associations: a test of optimal skew theory. Am. Nat. 145:119-132.
Reeve, H.K. 1997 Evolutionarily stable communication between kin: a general model. Proc. R. Soc. Lond. B 264:1037-1040.
Reilly, L.M. 1987 Measurements of inbreeding and average relatedness in a termite population. Amer. Natur. 130:339-349.
Rissing, S.W., G.B. Pollock, M.R. Higgins, R. H. Hager, and D.R. Smith 1989 Foraging specialization without relatedness or dominance mong co-founding ant queens. Nature 338:420-422.
Roeloffs, R. & S.E. Riechert 1988 Dispersal and genetic structure of the cooperative spider, Agelena consociata. Evolution 42:173-183.
Ross, K.G. and R.W. Matthews 1989 Population genetic structure and social evolution in the eusocial sphecid wasp, Microstigmus comes (Hymenoptera). Am. Nat. 134:574-598.
Seger, J. 1991 Cooperation and conflict in social insects. In: Behavioral Ecology: an evolutionary approach. Third ed. (ed. J.R. Krebs & N.B. Davies) pp. 338-373. Blackwell Scientific, Boston.
Stern, D.L. & W.A. Foster 1996 The evolution of soldiers in aphids. Biol. Rev. 71:27-79.
Strassmann, J.E., C.R. Hughes, D.C. Queller, S. Trilliazi, R. Cervo, S.K. Davis, & K.F.
Goodnight 1989 Genetic relatedness in primitively eusocial wasps. Nature 342:268-270.
Trivers, R.L. and H. Hare 1976 Haplodiploidy and the evolution of the social insects. Science 191:249-263.