Recognition
I. Definition of recognition:
Discrimination and identification of a target individual or group [or habitat]
from among a field of similar nontarget individuals or groups [or habitats]
II. Types of recognition and possible benefits of discrimination
A. Species (direct benefits)
B. Prey, predators, parasites (direct benefits)
C. Habitat (direct benefits)
D. Mate (benefits for reproduction)
1. sex recognition
2. mate quality
E. Individual (direct benefits via reciprocity, cooperation, social integration)
F. Kin (indirect benefits via kin selection, nepotism and direct benefits via
inbreeding avoidance)
G. Group membership (direct benefits of group membership)
H. Self versus nonself (immunity)
III. Process of recognition
1. the sender provides information about its identity in the form of a
signal

2. the receiver perceives the signal against background noise
3. the receiver compares the signal to a model or template

4. the receiver decides whether sender is / is not the target
5. the receiver takes some action based on its decision
III. Central problem of recognition is discrimination of target from nontarget
1. Example of mother bat deciding whether to nurse a calling infant bat in a large
creche -- a dichotomous recognition task
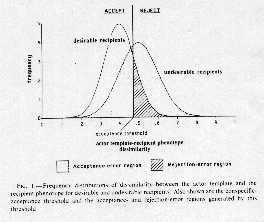
A. X axis describes the range of variation of the difference between the template
of the female bat and the actual phenotypes expressed by infant bats for both target
and nontarget classes (e.g. difference between a female bat's template of what her
offspring sounds like and the sounds that different classes of offspring in creche
make). Note that target and nontarget classes can have overlapping distributions.
B. If a mother bat has single cut-off in deciding to accept versus reject a calling
offspring, then then two types of error
a. Misses: missing giving aid to own offspring
b. False alarms: gives aid to nonrelated offspring
c. Moving cutoff can decrease one type of error but will increase other
d. Thus recognition is rarely perfect, and cheating is possible and may even be tolerated
2. How can this sort of relationship be changed through evolution?
A. Evolution of target either to make more distinct signals to minimize misses
or to make less distinct signals and take advantage of other mothers' false alarms
B. Evolution of receiver for increased discrimination ability -- e.g. take more
time, use more cues
3. In general, the more classes to be discriminated, the more complex the task involving
more memory and demanding more complex signals.
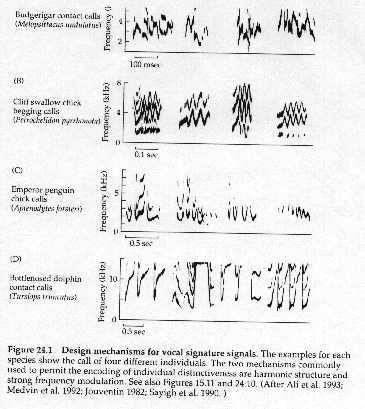
4. Most complex signals are required for individual recognition -- e.g. frequency
modulated signals with harmonic structure
IV. Mechanisms of recognition: three main classes, but many systems involve multiple
mechanisms
A. Spatial location -- recognition decision is based on proximity to a location
Examples: Brood parasitism in cowbirds, cuckoos, ducks -- parsitized birds raise
chicks as their own
Benefits: a simple decision rule, low decision costs, low cost to develop, no
selection on sender to develop special signal
Costs: easy to subvert by cheaters
B. Familiarity -- recognition is based on past experience followed by memorization
of specific characteristics; recognition decision is based on degree of matching
to this template
Example: Naked mole rats live in xenophobic colonies and treat unfamiliar animals,
kin or nonkin, with equal hostility; familiar or resident animals who are removed
from colony and reintroduced are accepted back into colony
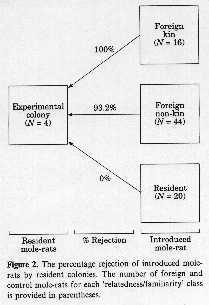
Example: Belding's ground squirrels -- use multiple recognition cues

Females chase close kin trespassers less often than distant kin or nonkin trespassers
suggesting that some kin recognition occurs
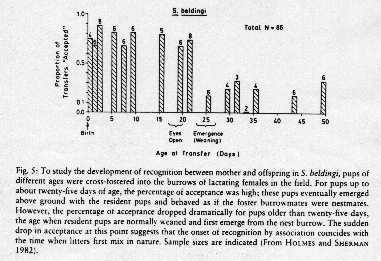
Up until age 25 days, pups can be cross fostered suggesting that spatial location
is an important recognition cue
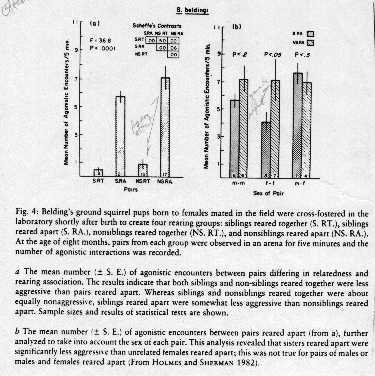
Sibs and nonsibs reared together treated with less aggression than sibs and nonsibs
reared apart (left graph) suggesting familiarity is important
For female-female pairs, sibs reared apart treated with less aggression than nonsibs
reared apart (right graph) suggesting yet another cue, not based on familiarity,
is used for kin recognition
Benefits: harder to cheat on that recognition based on spatial cues, allows greater
discriminability, can be used for individual, kin, or group level recognition
Costs: requires a variable signal, more memory
C. Phenotype matching -- functions to identify genetic similarity among close
kin
Uses a variable phenotype that is highly correlated to genetic similarity. The
receiver compares sender's phenotype to a know referent, usually either a familiar
relative or self
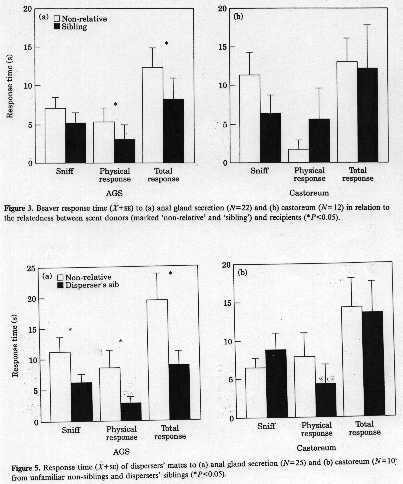
Example: Male beavers will scent mark on shores of bodies of water in their territories
and investigate the scent marks of other male beavers. They respond less strongly
to scent marks of siblings even though they have never come into direct contact with
those siblings (who are thus unfamiliar animals). This is true of scent marks based
on anogenital secretions (AGS) but not secretions from the castoreum. Interestingly,
mates of male beavers respond less strongly to the scent marks of their mates siblings
than to other males.
Example: MHC -- many recent studies support idea that MHC is used in kin recognition
is a variety of animal taxa
1.The major histocompatability complex is a set of hypervariable genetic loci
(many alleles) the products of which are cell surface protiens that seem to play
a major role in disease resistance, parasite detection, and recognition of self versus
nonself
2. Because of high allelic diversity, most individuals are heterozygotes and only
kin are likely to chare alleles
Experiments for MHC-based recognition
A) Create inbred mice that differed only at MHC locus
B) Allowed to associate in semi-natural enclosures in barns
C) Females preferred to mate with males whose MHC alleles differed from theirs
D) Females preferred to associate with MHC-similar males
E) Similar effects in shown in humans where females asked to rank the "pleasantness"
of smell of T-shirts worn by MHC-similar and MHC-different males
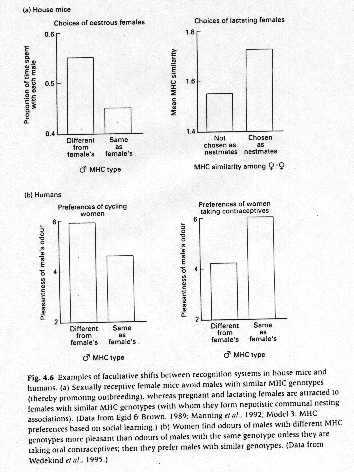
Benefits: mate with MHC-different to avoid inbreeding and associate with MHC-similar
for nepotism, kin selection, allows for recognition of close kin, very hard to subvert
Costs: may be difficult to evolve or to maintain suffient polymorphism in population
References:
Barnard, C. 1990 Kin recognition: problems, prospects and the evolution of discrimination
systems. Adv Stud Behav 19:29-81.
Beecher, M.D. 1989 Signalling systems for individual recognition: an information
theory approach. Anim. Behav. 38:248-261.
Beer, A.E., J.F. Quebbeman, J.W.T. Ayers, R.F. Haines 1981 Major histocompatibility
complex antigens, maternal and paternal immune responses, and chronic habitual abortions
in humans. Am. J. Obstet. & Gynecology 141:987-999.
Buss, L.W. 1982 Somatic cell parasitism and the evolution of somatic tissue compatibility.
Proc. Natl. Acad. Sci. 79:5337-5341.
Crozier, R.H. and M.E. Dix 1979 Analysis of two genetic models for innate components
of colony order in social hymenoptera. Behav. Ecol. Sociobiol. 4:217-224.
Crozier, R.H. 1986 Genetic clonal recognition abilities in marine invertebrates must
be maintained by selection for something else. Evolution 40:1100-1101.
Getz, W.M. 1981 Genetically based kin recognition systems. J. Theor. Biol. 92:209-226.
Grafen, A. 1990 Do animals really recognize kin? Anim. Behav. 39:42-54.
Greenberg, L. 1979 Genetic component of bee odor in kin recognition. Science 206:1095-1097.
Greenberg, L. 1988 Kin recognition in the sweat bee, Lasioglossum zephyrum. Behav.
Genet. 18:425-438.
Grosberg, R.K. and J.F. Quinn 1986 The genetic control and consequences of kin recognition
by the larvae of a colonial marine invertebrate. Nature 322:457-459.
Hamilton, W.D. 1964 The genetical evolution of social behavior. J. Theor. Biol. 7:1-52.
Hepper, P.G. 1991 Kin recognition. Cambridge University Press.
Lacy, R.C. and P.W. Sherman 1983 Kin recognition by phenotype matching. Am. Nat.
116:489-512.
Manning, C.J., Wakeland, E.K., and Potts, W.K. 1992 Communal nesting patterns in
mice implicate MHC genes in kin recognition. Nature 360:581-583.
Obin, M.S. and R.K. Vandermeer 1989 Mechanism of template-label matching in fire
ant, Solenopsis invicta Buren, nestmate recognition. Anim. Behav. 38:430-435.
Pfennig, D.W., P.W. Sherman, J.P. Collins 1994 Kin recognition and cannibalism in
polyphenic salamanders. Beh Ecol. 5:225-232.
Pfennig, D.W., P.W. Sherman 1995 Kin recognition. Sci. Am. 272:99-103.
Porter, R.H., S. A. McFadyen-Ketchum and G.A. King 1989 Underlying bases of recognition
signatures in spiny mice, Acomys cahirinus. Anim. Behav. 37:638-644.
Potts, W.K., Manning, C.J. & Wakeland, E.K. 1991 Mating patterns in seminatural
populations of mice influenced by MHC genotype. Nature 352:619-620.
Reeve, H.K. 1989 The evolution of conspecific acceptance thresholds. Amer. Natur.
133:407-435.
Sherman, P.W. and W.G. Holmes 1985 Kin recognition: issues and evidence. In: Experimental
Behavioral Ecology. (Holldobler, B. and M. Lindauer, eds) pp. 437-460, Sinauer, Sunderland.
Waldman, B. 1987 Mechanisms of kin recognition. J. Theor. Biol. 128:159-185.
Yamazaki, K., G.K. Beauchamp, C. Wysocki, J. Bard, L. Thomas and E.A. Boyse 1983
Recognition of H-2 types in relation to the blocking of pregnancy in mice. Science
221: 186-188.