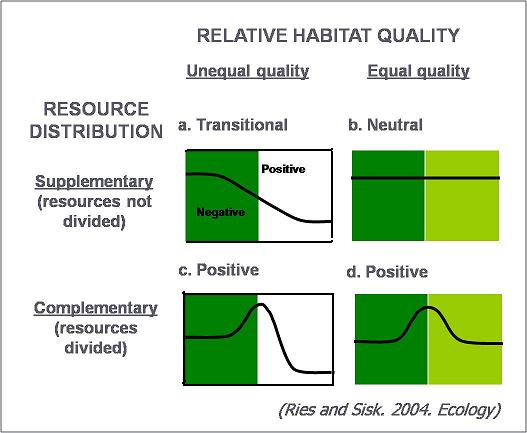
Our goal was to develop a model that was general enough that it could apply to any species on any landscape. Because of this, we focused on the two most basic mechanisms relating to edge responses: that the environment near the edge was likely to be a gradation from the conditions in the interior of one habitat to the interior of the other. Also, that if a species had resources that were divided between two habitats (the definition of complementary resources), that a positive edge effect is the most likely because being near the edge offers the best access to all necessary resources. This led to a conceptual model that divided responses into four categories based on two factors: relative habitat quality and resource distribution). When habitat quality was different on either side of the edge, we expect a transition in density across an entire edge gradient as species map onto the changes in habitat quality there. If habitat quality is equal, then no edge response is predicted. These dynamics are generated directly from mechanism 1, but also incorporate mechanism 3 (species "mapping"). Finally, if resources are complementary (divided), we always predict increased being near the edge (mechanism 2). This model does not incorporate changes in species interaction (mechanism 4) because we could not see a way to do that without considering species and system-specific dynamics that would necessarily make the model less general. Below, we show a simple representation of edge responses that would be predicted given different combinations of habitat quality and resource distribution.
Because this model is so simple, it can be applied to any species in any landscape. There are also some interesting implications that emerge from this conceptual framework:
1) All species are predicted to show positive, neutral, and negative edge responses, depending on the edge type encountered. This should move the ecological community away from the idea that species are intrinsically edge-loving or edge-avoiding (as also suggested by Villard, Imbeau, Baker
2) In order to understand observed edge responses, it is necessary to know the quality of the habitat on both sides of the edge. A review of the edge literature shows that many studies do not even describe what the adjacent habitat is composed of, or uses only vague descriptions like "open". This may explain much of the observed variability in the edge literature because responses would be compared between edge types that were different.
3) Finally, predictable changes in resource use (i.e., during different seasons or regions) may explain observed variability. To test this idea, it is necessary to compare responses when it is known that different spatial or temporal patterns exist in habitat use or resource distribution - and to date we have not found a good system or set of stufies to test this, but this should be a focus of future research.
We used the model to make predictions both based on literature reviews (Ries et al. 2004) and an extensive field project on butterflies in southeastern Arizona (Ries and Sisk 2008). The results suggest that when edge responses are observed, they are largely in the predicted direction. However, unpredicted neutral responses (where we expected a response and didn't observe one) was the most common form of unexplained variability. The below figure divides all responses (either culled from the literature or from our field work) into cases where observations compared to predictions were either correct, or incorrect. Incorrect outcomes were divided between cases where no response was observed when one was predicted ("neutral") or where an unpredicted edge response was observed ("wrong"). The top box shows the 3x3 contingency tables that describes all the different outcomes.
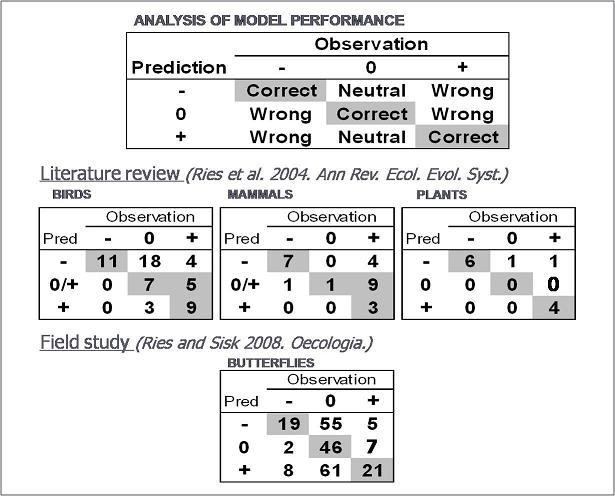
Another recent study that predicted bird edge responses based on past classifications of whether they were edge or interior species also resulted in many unpredicted neutral responses, but not observed responses in the opposite direction (Villard et al. 2007). Because of the prevalence of unpredicted neutral results, we have been developing the concept that certain species may be intrinsically insensitive to edges or that their sensitivity may vary depending on exrinsic conditions such as edge contrast (Ries and Sisk, 2010). The idea of edge sensitivity is potentially a very useful one, and we advocate that instead of categorizing species as to whether they are edge-avoiding or edge-loving (as has been commonly done in the past, but is really an artefact of a traditional focus on forest edges) it would be better to categorize species as to their sensitivity to edges. Further, we doubt that if edge sensitivity is variable, there is likely a continuum of response. Based on this, we advocate future research that ranks species according to their sensitivity to edges and then determines if their are any life history or ecological traits that are consistently related to sensitivity. We have used this approach to show that edge contrast does not seem to influence the likelihood of observing predicted edge responses in desert riparian butterflies, but that butterflies with lighter color wings and more vulnerable to predation seem more sensitive to the presence of edges (Ries and Sisk, 2010).
Other research that has drawn from this framework include a recent study was also done that used the model to predict community responses in seagrass beds (Macreadie et al. 2010). Lindell et al (2007) also used our framework to understand differential responses between temperate and tropical birds to forest edges. Mueller et al. (2009) showed that the complementarity described in the model can act differently at different scales.