Organismal Regulation
Given the numerous components of a cell, how do animals keep any two cells similar? This fundamental question has received little attention compared to unequal or asymmetric cell division. Yet, equal cell division is deployed repeatedly by multicellular organisms. During early development, all the information for making an organism can be divided equally as evidenced by the presence of monozygotic twins. Later, after different tissues have been specified through unequal cell divisions, equal cell divisions can begin to generate similar cells that populate a tissue. Thus, organisms must also have mechanisms that enable bidirectional switching between unequal and equal cell divisions.
Tissue Homogeneity: We have begun to reveal mechanisms that ensure effective switching from unequal to equal cell division. We found that expression from repetitive DNA is particularly vulnerable to variation between cells. 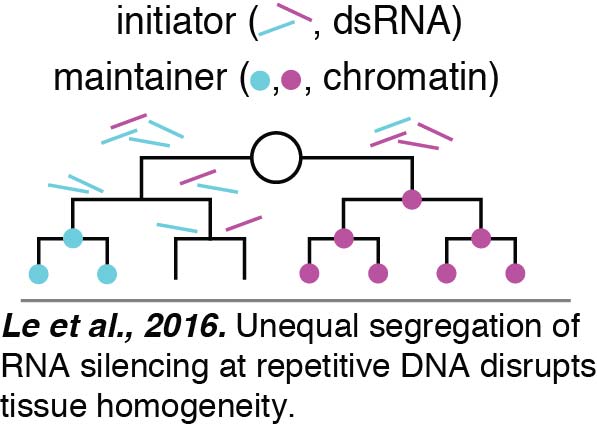 Our analyses suggests that this variation between cells results from unequal RNA silencing. An inhibitor of RNA silencing, ERI-1, that is inherited from parents as well as made in developing progeny acts in the embryo to eliminate such variation between cells and ensure uniform expression. In the absence of this factor, an initiator of gene silencing, likely double-stranded RNA (dsRNA), is segregated unequally between cells during the proliferative cell divisions required to make a tissue. This initiator causes threshold-dependent gene silencing. Consistent with the initiator being dsRNA, fewer cells show silencing when the dsRNA importer SID-1 is removed to block the spread of dsRNA between cells. Once initiated, the silencing is maintained by chromatin-mediated mechanisms despite DNA replication and cell division (Le et al., 2016).
Our analyses suggests that this variation between cells results from unequal RNA silencing. An inhibitor of RNA silencing, ERI-1, that is inherited from parents as well as made in developing progeny acts in the embryo to eliminate such variation between cells and ensure uniform expression. In the absence of this factor, an initiator of gene silencing, likely double-stranded RNA (dsRNA), is segregated unequally between cells during the proliferative cell divisions required to make a tissue. This initiator causes threshold-dependent gene silencing. Consistent with the initiator being dsRNA, fewer cells show silencing when the dsRNA importer SID-1 is removed to block the spread of dsRNA between cells. Once initiated, the silencing is maintained by chromatin-mediated mechanisms despite DNA replication and cell division (Le et al., 2016).
Once different tissues have been specified through unequal cell divisions and signaling in the early embryo, subsequent cell divisions within a tissue need to generate similar cells. Given the fundamental nature of this requirement, we suspect that many more processes in cells are under similar control to ensure a clean switch from differentiation to epigenetic memory during proliferative cell divisions that generate a tissue. Loss of such developmental mechanisms could potentially cause diseases later in life because of the generation of a variable population of cells within a tissue. For example, such pre-existing variation could increase the odds of age-related disease such as cancer that strike single cells apparently at random.
Functional Mosaicism: We found that cells can appear similarly functional despite underlying differences that vary from animal to animal. 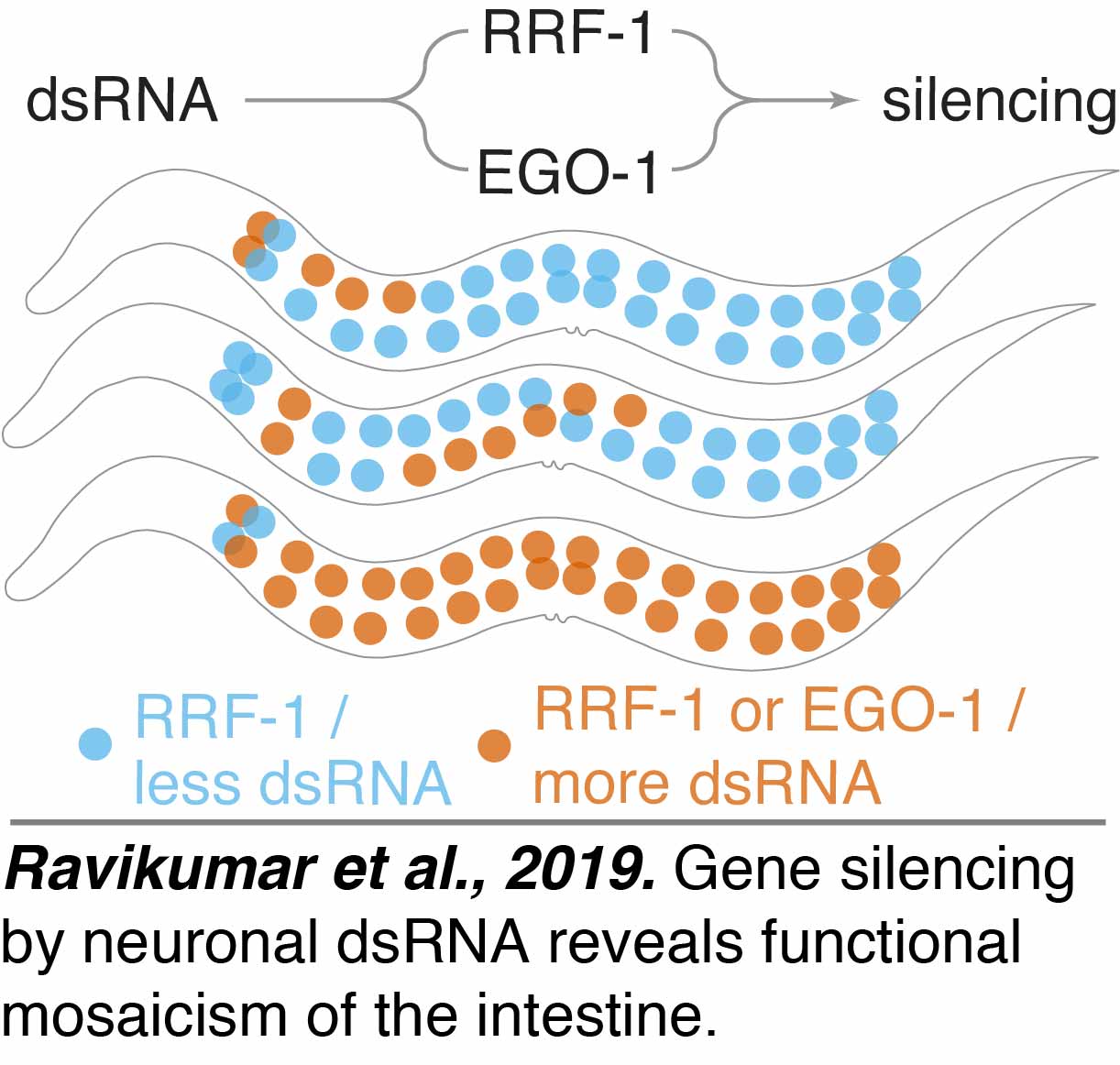 Early asymmetric cell divisions separate the germline from the soma in C. elegans and it was thought that RNA interference relied on small RNAs amplified using two 'tissue-specific' RNA-dependent RNA polymerases (RdRPs) - EGO-1 in the germline and RRF-1 in somatic cells. We found that EGO-1 can compensate for the lack of RRF-1 when dsRNA from neurons is used to silence genes in intestinal cells. However, the lineal identity of cells that can use EGO-1 varies (Ravikumar et al., 2019). This functional mosaicism or equivalence that allows for underlying mosaicism cautions against inferring uniformity of mechanism based on uniformity of outcome.
Early asymmetric cell divisions separate the germline from the soma in C. elegans and it was thought that RNA interference relied on small RNAs amplified using two 'tissue-specific' RNA-dependent RNA polymerases (RdRPs) - EGO-1 in the germline and RRF-1 in somatic cells. We found that EGO-1 can compensate for the lack of RRF-1 when dsRNA from neurons is used to silence genes in intestinal cells. However, the lineal identity of cells that can use EGO-1 varies (Ravikumar et al., 2019). This functional mosaicism or equivalence that allows for underlying mosaicism cautions against inferring uniformity of mechanism based on uniformity of outcome.
We speculate that functional mosaicism and its control could be common in multicellular organisms because of the need to balance diversification of cell types with preservation of fundamental functions in all cells. This property of organisms could contribute to escape from targeted therapies and could allow developmental systems to drift over evolutionary time.
Stochastic germline expression: We found that the two gonad arms of the C. elegans germline can show dramatic differences in expression. 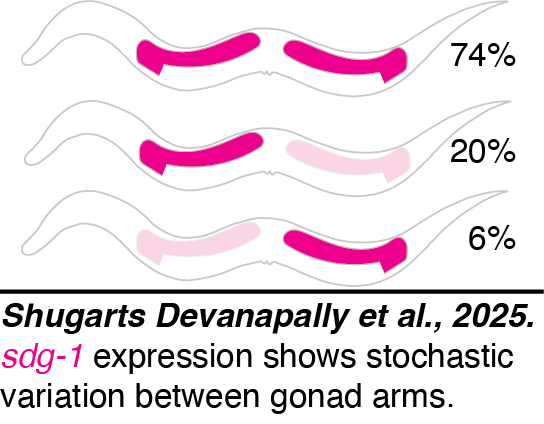 Expression of the SID-1-dependent gene sdg-1 can randomly vary such that either gonad arm can show reduced expression (Shugarts Devanapally et al., 2025) and yet expression remains stable across generations. One hypothesis is that during the independent development of each gonad arm after descendants of Z2/Z3 cells separate, differences in RNA silencing arise but are insufficient for causing heritable RNA silencing. The mechanisms that mediate this silencing and the evolutionarily selected function, if any, for the regulation of such variation is unclear. Additional experiments are required to understand the basis of this stochastic variation in gene expression within the germline.
Expression of the SID-1-dependent gene sdg-1 can randomly vary such that either gonad arm can show reduced expression (Shugarts Devanapally et al., 2025) and yet expression remains stable across generations. One hypothesis is that during the independent development of each gonad arm after descendants of Z2/Z3 cells separate, differences in RNA silencing arise but are insufficient for causing heritable RNA silencing. The mechanisms that mediate this silencing and the evolutionarily selected function, if any, for the regulation of such variation is unclear. Additional experiments are required to understand the basis of this stochastic variation in gene expression within the germline.
Back to research